
| 时间:2024-08-30 |
2024年发表于《Nucleic Acids Research》(IF=16.6)
一、研究背景
当环境条件发生剧烈和不利变化时,细胞会通过应激反应基因的转录重编程来调节RNA合成以适应变化。RNA结合蛋白(RBP)参与转录后调控,最近发现有一部分RBP在开放的染色质区域,如增强子和启动子,称为染色质相互作用RBP(ChRBP)。ChRBP的上游调控因子及其机制性质仍然具有挑战性和悬而未决的问题。circRNA(circRNAs)是一类单链和共价封闭转录本,可能调控染色质上ChRBP分布和功能活动,相关研究仍然非常有限。
二、研究结果
1、一类与gawky相互作用的circRNA的鉴定
假设某些circRNAs可能在ChRBP gawky蛋白介导的转录激活中充当上游调节因子,FLAGgawky稳定细胞系,铜应激细胞进行CLIP-seq(图1A)。19个circRNA与铜激活的gawky高度相关(图1B, C)。使用FIRE软件,发现MRE基序在gawky相互作用的circRNAs的RNA序列中比打乱的序列控制更丰富(图1D)。DEAD-box RNA解旋酶Hlc可以特异性地维持应激反应基因的基础转录,而不是应激诱导的转录,FLAGHlc细胞的CLIP-seq表明FLAG-Hlc对MRE circRNA的结合能力非常有限。MRE circRNAs不与非胁迫的gawky或胁迫的Hlc结合(图1G),MRE circRNA-gawky相互作用的特异性。MRE circRNA同时存在于细胞核和细胞质部分(图1I)。表明MRE circRNA是一类应力响应型circRNA,在金属胁迫下特异性地与gawky相互作用。
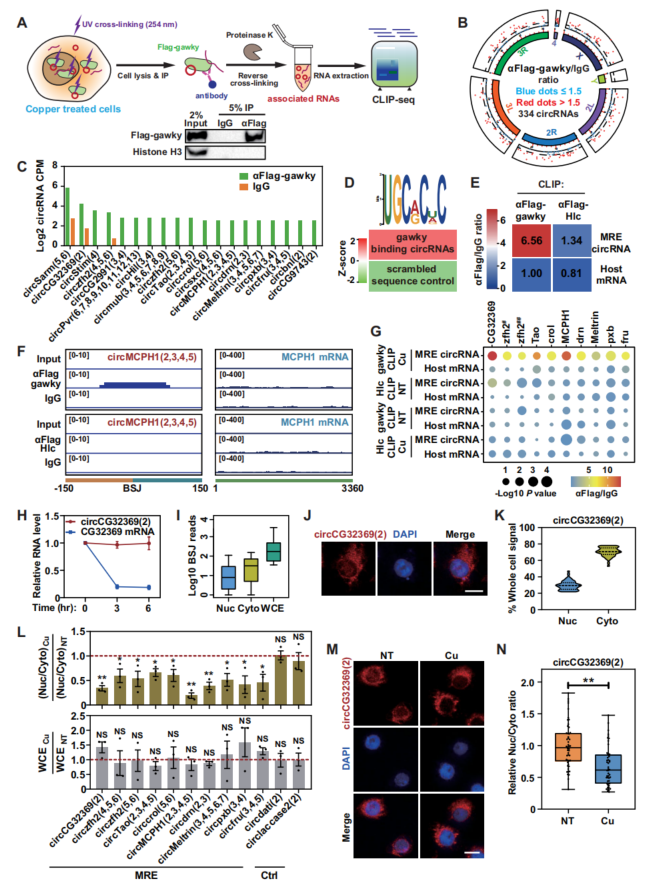
图1 MRE circRNA的鉴定
2、MRE circRNA可以减弱应激诱导的转录
用circCG32369和circMCPH1载体构建MRE circRNA稳定细胞系(图2A),并通过RT-qPCR确定过表达circCG32369或circMCPH1(图2B)。与正常的S2细胞相比,在铜胁迫下过表达MRE circRNAs的细胞,MT基因(一种特征明确的金属胁迫诱导基因)的mRNA水平被大幅下调(图2C)。在不含MRE的对照circRNA过表达时,MT的表达基本保持不变(图2F、G)。线性RNA稳定的细胞系中,应力诱导的MT表达没有改变(图2F, G),可知MRE circRNA的线性状态不会导致MT下调。因此,这些数据排除了MRE circRNA可能通过调节其稳定性或核输出来调节MT表达的可能性。
NRO实验研究circCG32369和circMCPH1是否调节MT的转录活性,发现MT mRNA的新生转录物丰度显著降低(图2I),支持MRE circRNA(circCG32369和circMCPH1),可以作为有效的转录抑制剂响应铜胁迫。MRE circRNA敲除证实,单个MRE circRNA的功能丧失不足以激发MT转录。
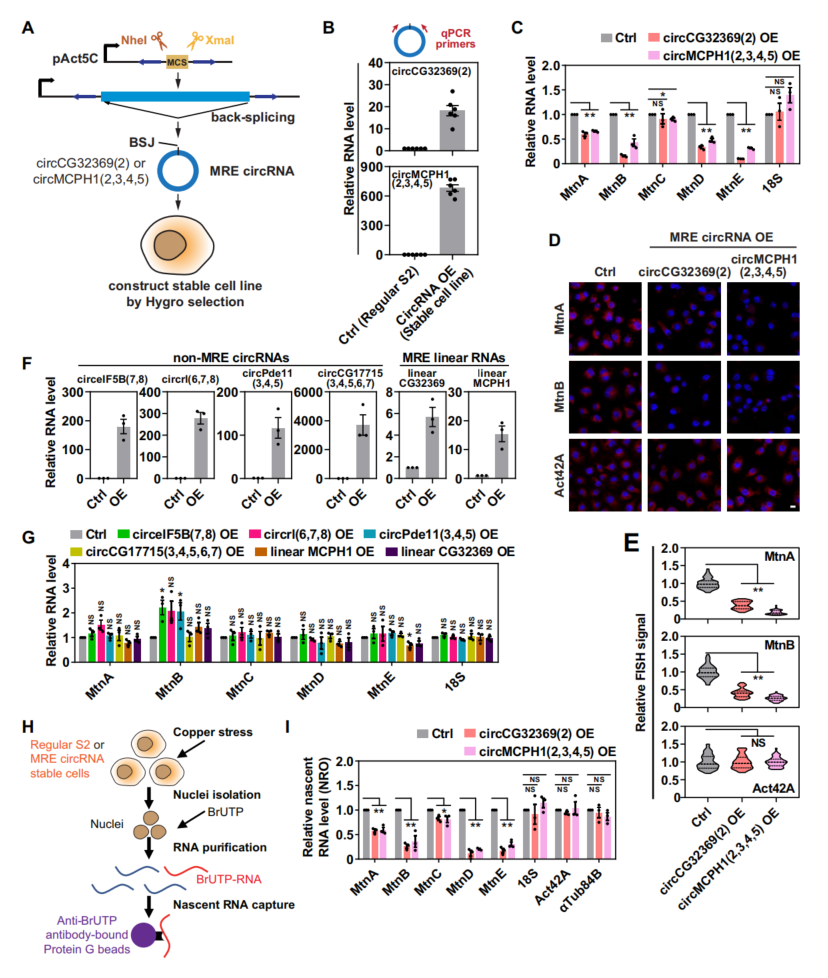
图2 MRE circRNA是应激诱导转录的有效抑制剂
3、MRE circRNA的功能活性需要gawky的存在
为了验证MRE circRNA介导的转录抑制是否在启动子区域是特异性结合,使用外显子mini-dati基因载体,加上MT启动子,并将其转入MRE circRNA稳定细胞中。硫酸铜胁迫细胞后,通过RT-qPCR发现,在两个MRE circRNA稳定细胞系中,mini-dati基因的表达水平被显著抑制,这与内源性MT的表型相似(图2C)。表明,MRE circRNA在转录中的准确作用依赖于启动子上下游。前述研究已知MRE circRNA特异性地与gawky相互作用(图1A-G,gawky通过启动子调控金属响应基因的转录激活,因此推断MRE circRNAs和gawky可能在一定程度上功能偶联。MRE circRNAs的过表达导致β-gal dsRNA(对照dsRNA)处理的细胞中MtnB mRNA水平显著降低(图3E)。相比之下,MRE circRNA无法抑制gawky-depletion细胞中的MtnB转录(图3E)。同样,在没有gawky的情况下,MRE circRNAs的过表达不再影响Pol II与MtnB的结合(图3F)。综上所述,gawky的存在是MRE circRNA抑制应激诱导转录的先决条件。
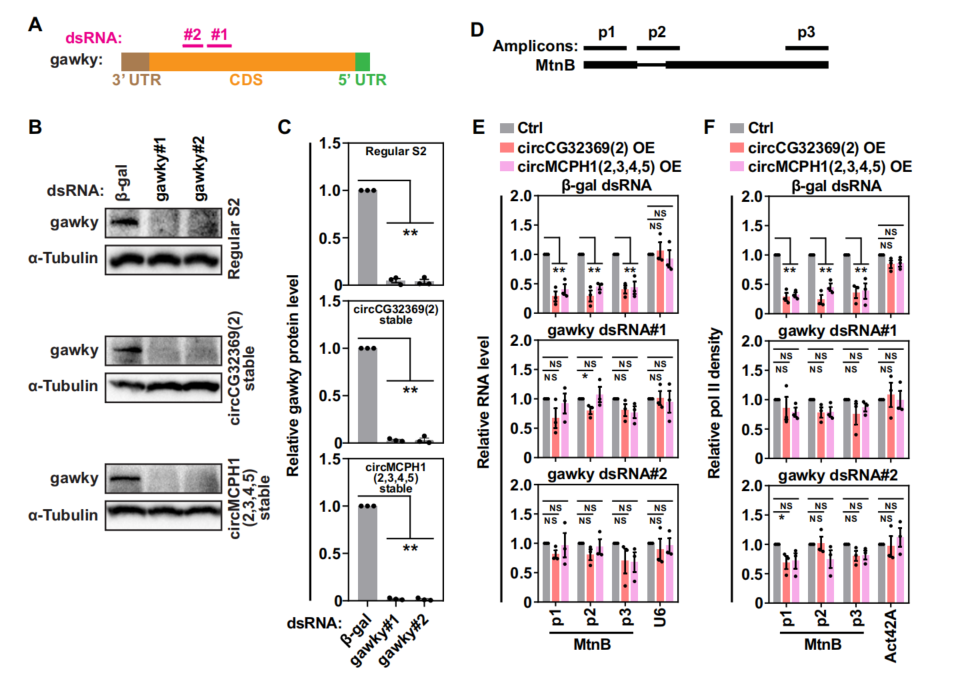
图3 MRE circRNA在gawky介导的转录中起作用
4、MRE circRNA调节gawky的分子作用
通过内源性gawky的抗体进行ChIP-seq,发现gawky与MtnB启动子之间存在强相互作用(图4A,灰线)。在稳定过表达MRE circRNAs的细胞中,gawky ChIP信号急剧减少(图4A,粉色和紫色线),尽管检测到的gawky总表达水平没有显著差异(图4B、C)。表明MRE circRNAs损害了gawky向活性基因位点的募集。结合MTF-1 ChIP结果,证实了MRE circRNA介导的调控在胁迫诱导转录中的特异性。免疫荧光染色分析显示,在铜胁迫条件下,MRE circRNA可能在某种程度上作为gawky载体,有效地输出gawky和/或阻止gawky进入细胞核。综上所述,MRE circRNAs通过调节gawky的分子作用(即从应激反应基因中释放gawky并增强其细胞质积累)特异性地调节应激诱导的转录。
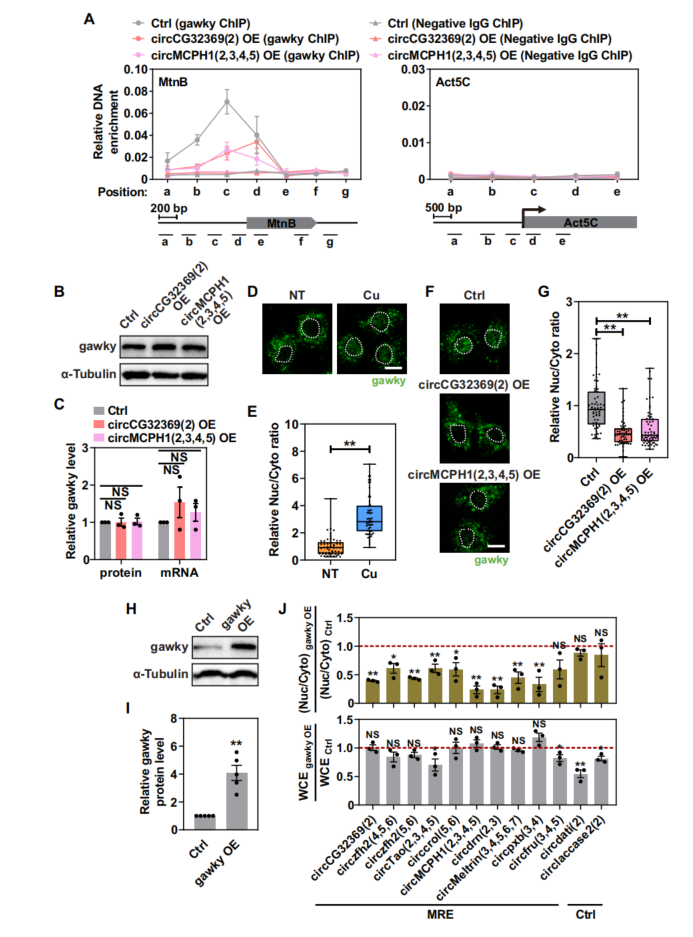
图4 MRE circRNA调节gawky的分子作用
5、MRE基序对于MRE circRNA在转录抑制中的准确功能具有重要意义
生成突变circCG32369载体构建稳定细胞系(图5A),进行gawky CLIP-seq,与野生型相比,circCG32369突变体在铜胁迫下仅表现出有限的对gawky的结合能力(图5B);且不是MRE突变后circCG32369丰度改变的结果(图5C)。发现表明circCG32369-gawky相互作用依赖于位于circRNA中的MREs。gawky的亚细胞定位,发现circCG32369在其MRE突变时诱导应激激活gawky的细胞质积累的能力大大受损(图5G、H)。因此,在MREs缺失后,circCG32369对铜诱导的MtnB转录的抑制作用减弱(图5I)。综上所述,数据证明位于MRE circRNAs中的MREs是circRNA-gawky相互作用的调控元件,这对于在应激条件下精确调控gawky介导的转录是必不可少的。
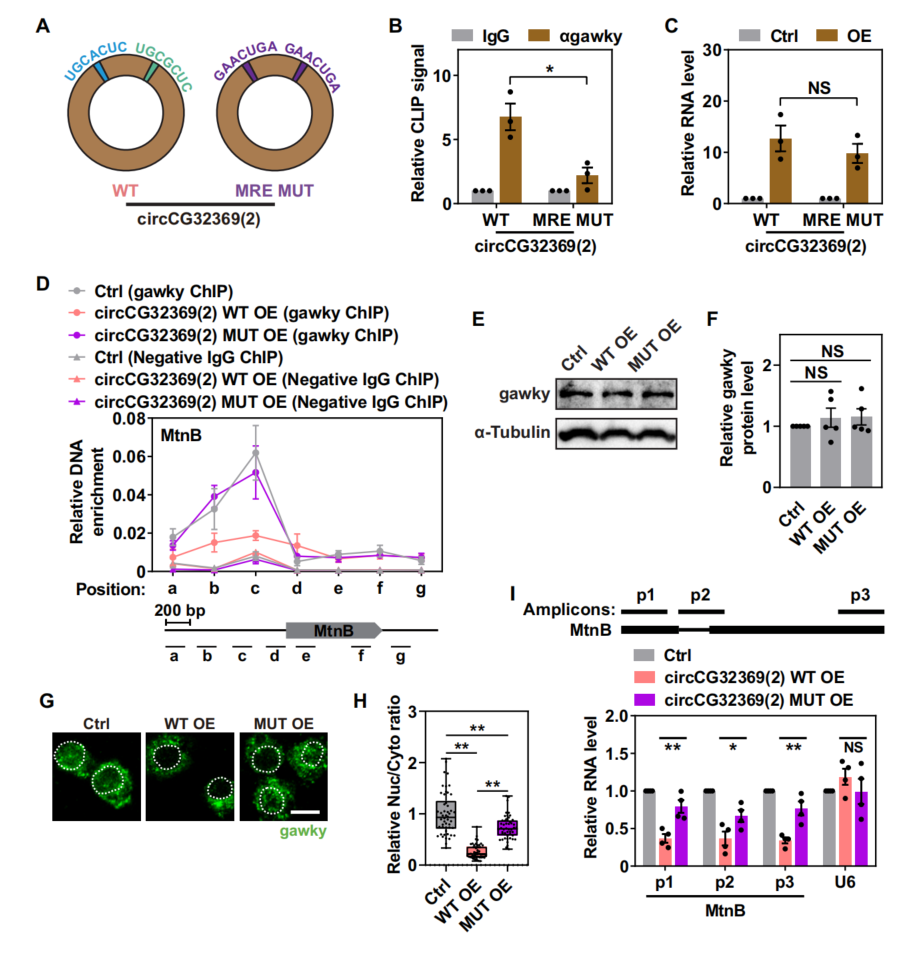
图5 MRE基序是MRE circRNA介导的转录抑制所必需的
6、铜胁迫时,MRE circRNA对数百个铜诱导基因有很强的调控能力
硫酸铜处理后,共有1627个基因的表达水平显著升高(图6A),称为铜诱导基因。circCG32369过表达导致71.3%的差异表达基因被显著抑制(图6A),具有强烈的全局抑制作用(图6B),这意味着MRE circRNA对铜诱导基因群具有选择性。铜诱导基因对circCG32369介导的转录调控更为敏感,circCG32369更倾向于沉默诱导水平较高的基因(图6F、G)。高表达基因对circCG32369过表达的敏感性较低(图6N、O)。综上所述,MRE circRNA在抑制应激诱导转录中的广泛作用,并清楚地揭示了它们对应激诱导基因的选择性作用。敲低单个MRE circRNA不能促进应激诱导的转录,两种MRE circRNA对某些基因群具有明显的选择性。证明了不同的MRE circRNA对不同基因的转录重编程的组合控制。
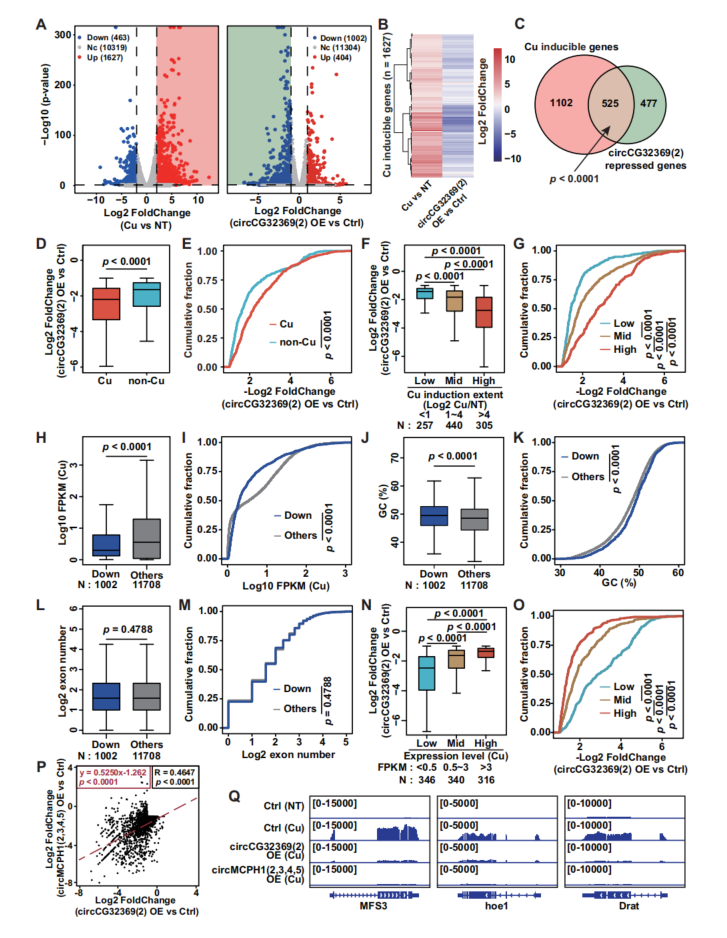
图6 MRE circRNA对应激诱导转录的抑制作用在许多果蝇基因中普遍存在
7、铜胁迫下MRE circRNA与细胞内稳态的生理相关性
铜诱导细胞毒性的最重要的一种机制,由铜离子还原而成的亚铜离子能催化羟基自由基的生成,可进一步导致氧化损伤和DNA链断裂。RNA-seq显示一系列抗氧化防御和DNA损伤修复相关基因在细胞对铜胁迫的防御中显着上调(图7A)。在铜应激细胞的细胞核中显示出明显的DNA损伤信号(图7B、C)。与常规的S2细胞对照相比,所有检测到的抗氧化防御和DNA损伤修复相关基因在两种MRE circRNA稳定细胞系中都被显著抑制(图7A);MRE circRNAs的过表达显著加重了铜诱导的DNA损伤(图7B, C)。MRE circRNAs的过表达显著降低了细胞的铜抗性(图7D)。综上所述,这些发现证实了MRE circRNA在铜胁迫下促进受损细胞清除的生理相关性。
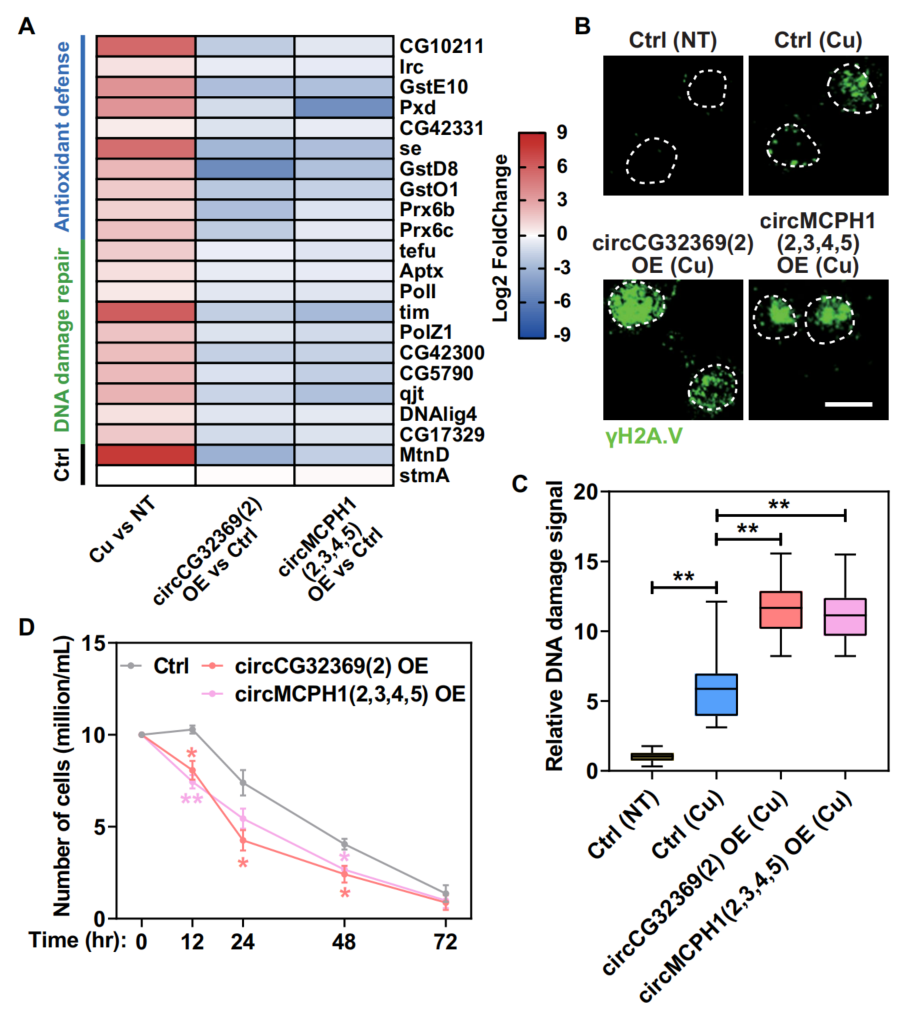
图7 MRE circRNA加重了铜诱导的DNA损伤
三、研究结论
在重金属胁迫下,与gawky蛋白相互作用的MRE circRNA可拮抗gawky在活性染色质区域的结合,并诱导其异常的细胞质积累来抑制gawky的功能活性,特异性地调节应激诱导的转录。circRNAs中的MRE基序被确定为维持circRNA-gawky相互作用的必要RNA调控因子。circRNA-RBP染色质轴可能是真核细胞中基因表达的基本调控网络。
参考文献:
A circular RNA-gawky-chromatin regulatory axis modulates stress-induced transcription. [J]Nucleic Acids Research, 2024.